生物化学与分子生物学/乳糖操纵元的表达调控
| 医学电子书 >> 《生物化学与分子生物学》 >> 基因表达调控 >> 原核基因表达调控 >> 乳糖操纵元的表达调控 |
| 生物化学与分子生物学 |
|
|
|
如上所述乳糖操纵元的结构及其基因表达调控可综合于图19-7。
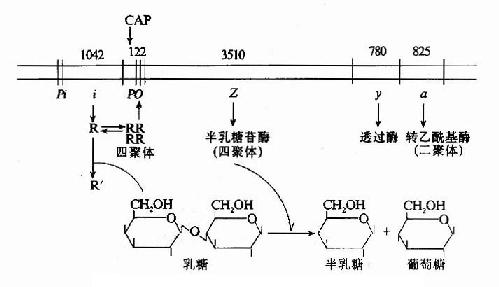
图19-7 乳糖操纵元的结构及调控示意图
(一)阻遏蛋白的负性调控
当大肠杆菌在没有乳糖的环境中生存时,1ac操纵元处于阻遏状态。i基因在其自身的启动子Pi控制下,低水平、组成性表达产生阻遏蛋白R,每个细胞中仅维持约10个分子的阻遏蛋白。R以四聚体形式与操纵子o结合,阻碍了RNA聚合酶与启动子P1ac的结合,阻止了基因的转录起动。R的阻遏作用不是绝对的,R与o偶尔解离,使细胞中还有极低水平的β-半乳糖苷酶及透过酶的生成。
当有乳糖存在时,乳糖受β-半乳糖苷酶的催化转变为别乳糖,与R结合,使R构象变化,R四聚体解聚成单体,失去与o的亲和力,与o解离,基因转录开放,β-半乳糖苷酶在细胞内的含量可增加1000倍。这就是乳糖对1ac操纵元的诱导作用。
一些化学合成的乳糖类似物,不受β-半乳糖苷酶的催化分解,却也能与R特异性结合,使R构象变化,诱导1ac操纵元的开放。例如异丙基硫代半乳糖苷(isopropylthiogalactoside,IPTG)就是很强的诱导剂,不被细胞代谢而十分稳定。X-gal(5-溴-4-氯-3-吲哚-β-半乳糖苷)也是一种人工化学合成的半乳糖苷,可被β-半乳糖苷酶水解产生兰色化合物,因此可以用作β-半乳糖苷酶活性的指示剂。IPTG和Xgal都被广泛应用在分子生物学和基因工程的工作中。
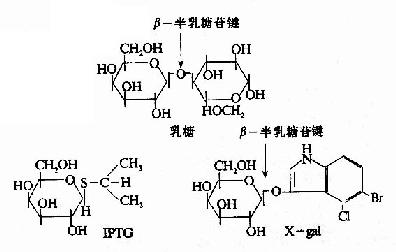
图19-8 乳糖,IPTG和Xgal的结构
(二)CAP的正性调控
细菌中的cAMP含量与葡萄糖的分解代谢有关,当细菌利用葡萄糖分解供给能量时,cAMP生成少而分解多,cAMP含量低;相反,当环境中无葡萄糖可供利用时,cAMP含量就升高。细菌中有一种能与cAMP特异结合的cAMP受体蛋白CRP(cAMp receptor protein),当CRP未与cAMP结合时它是没有活性的,当cAMP浓度升高时,CRP与cAMP结合并发生空间构象的变化而活化,称为CAP(CRPcAMp activated protein),能以二聚体的方式与特定的DNA序列结合。
在1ac操纵元的启动子P1ac上游端有一段与Plac部分重叠的序列,能与CAP特异结合,称为CAP结合位点(CApbinding site)。CAP与这段序列结合时,可增强RNA聚合酶的转录活性,使转录提高50倍。相反,当有葡萄糖可供分解利用时,cAMP浓度降低,CRP不能被活化,1ac操纵元的结构基因表达下降。
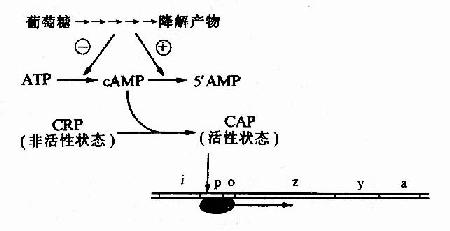
图19-9 葡萄糖利用对乳糖操纵元的影响
由于P1ac是弱启动子,单纯因乳糖的存在发生去阻遏使1ac操纵元转录开放,还不能使细胞很好利用乳糖,必须同时有CAP来加强转录活性,细菌才能合成足够的酶来利用乳糖。1ac操纵元的强诱导既需要有乳糖的存在,又需要没有葡萄糖可供利用。通过这种机制,细菌优先利用环境中的葡萄糖,只有无葡萄糖而又有乳糖时,细菌才去充分利用乳糖。
细菌对葡萄糖以外的其他糖(如阿拉伯糖、半乳糖、麦芽糖等)的利用上也有类似对乳糖利的情况,在含有编码利用阿拉伯糖的酶类基因群的阿拉伯糖操纵元(araoperon)、半乳糖操纵元(gal operon)中也有CAP结合位点,CAP也起类似的正性调控作用。所以CAP的通用名称是分解代谢基因激活蛋白(catabolic gene activator protein)。
不难看出:CAP结合位点就是一种起正性调控作用的操纵子,CAP则是对转录起正性作用的控蛋白棗激活蛋白,编码CRP的基因也是一个调控基因,不过它并不在1ac操纵元的附近,CAP可以对几个操纵元都起作用。
从上所述,乳糖操纵元属于可诱导操纵元(inducible operon),这类操纵元通常是关闭的,当受效应物作用后诱导开放转录。这类操纵元使细菌能适应环境的变化,最有效地利用环境能提供的能源底物。
| 关于“生物化学与分子生物学/乳糖操纵元的表达调控”的留言: | |
|
目前暂无留言 | |
| 添加留言 | |