生物化学与分子生物学/蛋白质生物合成过程
| 医学电子书 >> 《生物化学与分子生物学》 >> 蛋白质的生物合成 >> 蛋白质生物合成过程 |
| 生物化学与分子生物学 |
|
|
|
蛋白质生的合成亦称为翻译(Translation),即把mRNA分子中碱基排列顺序转变为蛋折质或多肽链中的氨基酸排列顺序过程。这也是基因表达的第二步,产生基因产物蛋白质的最后节段。不同的组织细胞具有不同的生理功能,是因为它们表达不同的基因,产生具有特殊功能的蛋白质,参与蛋白质生物合成的成份至少有200种,其主要体第主要由mRNA、tRNA、核糖核蛋白体以及有关的酶和蛋白质因子共同组成。
原核生物与真核生物的蛋白质合成过程中有很多的区别,真核生物此过程更复杂,下面着重介绍原核生物蛋白质合成的过程,并指出真核生物与其不同这处。
蛋白质生物合成可分为五个阶段,氨基酸的活化、多肽链合成的起始、肽链的延长、肽链的终止和释放、蛋白质合成后的加工修饰。
目录 |
(一)氨基酰-tRNA的生成
氨基酸在进行合成多肽链之前,必须先经过活化,然后再与其特异的tRNA结合,带到mRNA相应的位置上,这个过程靠氨基酰tRNA合成酶催化,此酶催化特定的氨基酸与特异的tRNA相结合,生成各种氨基酰tRNA.每种氨基酸都靠其特有合成酶催化,使之和相对应的tRNA结合,在氨基酰tRNA合成酶催化下,利用ATP供能,在氨基酸羧基上进行活化,形成氨基酰-AMP,再与氨基酰tRNA合成酶结合形成三联复合物,此复合物再与特异的tRNA作用,将氨基酰转移到tRNA的氨基酸臂(即3'-末端CCA-OH)上(图18-5和图18-6)。
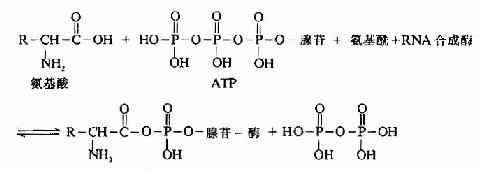
图18-6 氨基酰-tRNA的生成
原核细胞中起始氨基酸活化后,还要甲酰化,形成甲酰蛋氨酸tRNA,由N10甲酰四氢叶酸提供甲酰基。而真核细胞没有此过程。
前面讲过运载同一种氨基酸的一组不同tRNA称为同功tRNA。一组同功tRNA由同一种氨酰基tRNA合成酶催化。氨基酰tRNA合成酶对tRNA和氨基酸两者具有专一性,它对氨基酸的识别特异性很高,而对tRNA识别的特异性较低。
氨基酰tRNA合成酶是如何选择正确的氨基酸和tRNA呢?按照一般原理,酶和底物的正确结合是由二者相嵌的几何形状所决定的,只有适合的氨基酸和适合的tRNA进入合成酶的相应位点,才能合成正确的氨酰基tRNA。现在已经知道合成酶与L形tRNA的内侧面结合,结合点包括接近臂,DHU臂和反密码子臂(图18-7)。
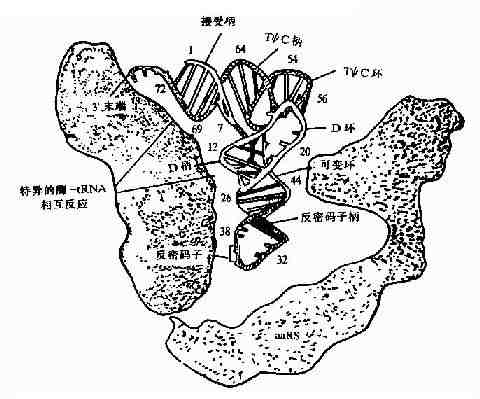
图18-7 氨基酰-tRNA合成酶与tRNA的相互作用,可见氨酸接受柄、
D柄、反密码子和可变环与酶反应
乍看起来,反密码子似乎应该与氨基酸的正确负载有关,对于某些tRNA也确实如此,然而对于大多数tRNA来说,情况并非如此,人们早就知道,当某些tRNA上的反密码子突变后,但它们所携带的氨工酸却没有改变。1988年,候稚明和Schimmel的实验证明丙氨酸tRNA酸分子的氨基酸臂上G3:U70这两个碱基发生突变时则影响到丙氨酰tRNA合成酶的正确识别,说明G3:U70是丙氨酸tRNA分子决定其本质的主要因素。tRNA分子上决定其携带氨基酸的区域叫做副密码子。一种氨基酰tRNA合成酶可以识别以一组同功tRNA,这说明它们具有共同特征。例如三种丙氨酸tRNA(tRNAAlm/CUA,tRNAAim/GGC,tRNAAin/UGC都具有G3:U70副密码子。)但没有充分的证据说明其它氨基酰tRNA合成酶也识别同功tRNA组中相同的副密码子。另外副密码子也没有固定的位置,也可能并不止一个碱基对。
(二)多肽链合成的起始
核蛋白体大小亚基,mRNA起始tRNA和起始因子共同参与肽链合成的起始。
(1)核糖体30S小亚基附着于mRNA起始信号部位:原核生物中每一个mRNA都具有其核糖体结合位点,它是位于AUG上游8-13个核苷酸处的一个短片段叫做SD序列。这段序列正好与30S小亚基中的16s rRNA3’端一部分序列互补,因此SD序列也叫做核糖体结合序列,这种互补就意味着核糖体能选择mRNA上AUG的正确位置来起始肽链的合成,该结合反应由起始因子3(IF-3)介导,另外IF-1促进IF-3与小亚基的结合,故先形成IF3-30S亚基-mRNA三元复合物。
(2)30S前起始复合物的形成:在起始因子2作用下,甲酰蛋氨酰起始tRNA与mRNA分子中的AUG相结合,即密码子与反密码子配对,同时IF3从三元复合物中脱落,形成30S前起始复合物,即IF2-3S亚基-mRNA-fMet-tRNAfmet复合物,此步需要GTP和Mg2+参与。
(3)70S起始复合物的形成:50S亚基上述的30S前起始复合物结合,同时IF2脱落,形成70S起始复合物,即30S亚基-mRNA-50S亚基-mRNA-fMet-tRNAfmet复合物。此时fMet-tRNAfmet占据着50S亚基的肽酰位。而A位则空着有待于对应mRNA中第二个密码的相应氨基酰tRNA进入,从而进入延长阶段,以上过程见图18-8和图18-9。

图18-8 大肠杆菌起始复合物的形成
2、真核细胞蛋白质合成的起始
真核细胞蛋白质合成起始复合物的形成中需要更多的起始因子参与,因此起始过程也更复杂。
(1)需要特异的起始tRNA即,-tRNAfmet,并且不需要N端甲酰化。已发现的真核起始因子有近10种(eukaryote Initiation factor,eIF)
(2)起始复合物形成在mRNA5’端AUG上游的帽子结构,(除某些病毒mRNA外)
(3)ATP水解为ADP供给mRNA结合所需要的能量。真核细胞起始复合物的形成过程是:翻译起始也是由eIF-3结合在40S小亚基上而促进80S核糖体解离出60S大亚基开始,同时eIF-2在辅eIF-2作用下,与Met-tRNAfmet及GTP结合,再通过eIF-3及eIF-4C的作用,先结合到40S小亚基,然后再与mRNA结合。
mRNA结合到40S小亚基时,除了eIF-3参加外,还需要eIF-1、eIF-4A及eIF-4B并由ATP小解为ADP及Pi来供能,通过帽结合因子与mRNA的帽结合而转移到小亚基上。但是在mRNA5’端并未发现能与小亚基18SRNA配对的S-D序列。目前认为通过帽结合后,mRNA在小亚基上向下游移动而进行扫描,可使mRNA上的起始密码AUG在Met-tRNAfmet的反密码位置固定下来,进行翻译起始。
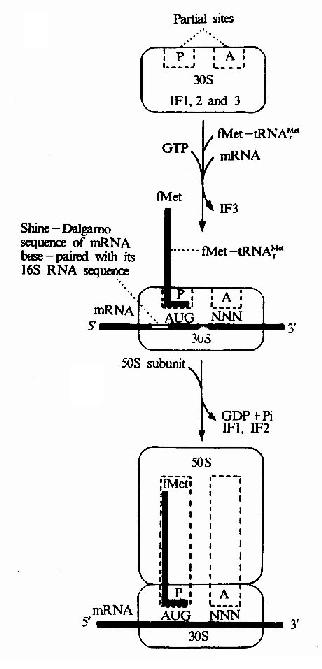
图18-9 Initiationof translation im E.cole.The initiating tRNA,tRNAMetf,is represented by theblue line,the anticodon being the horizontal short line.The fMet-tRNAMetf isdelivered to the 30s subunit by IF2.NNN represents any codon (N for anynucleotie).Note.The ribosome also has an exit site not shown in thediagram.This site will be discussed later.
通过eIF-5的作用,可使结合Met-tRNAfmet.GTP及mRNAR40S小亚基与60S大亚基结合,形成80S复合物。eIF-5具有GTP酶活性,催化GTP水解为GDP及Pi,并有利于其它起始因子从40S小亚基表面脱落,从而有利于40S与60S两个亚基结合起来,最后经eIF-4D激活而成为具有活性的80SMet-tRNAfmet.mRNA起始复合物。
真核细胞翻译起始复合物的生成见图18-10和图18-11。
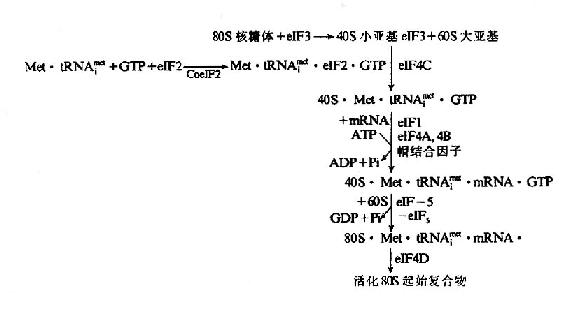
图18-10 真核细胞翻译起始复合物的形成
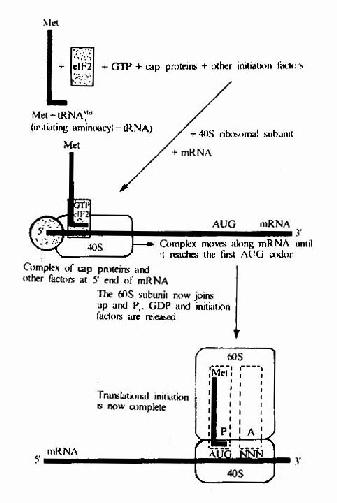
图18-11 Simplified diagram of initiation in eukaryotes.Note that severaleukaryote initiation factors besides elF2 are involved .tRNAMeti,initiatingRNA.e1F2 is the eukaryotic initiation factor corresponding to IF2 inprokaryotes.
(三)多肽链的延长:
在多肽链上每增加一个氨基酸都需要经过进位,转肽和移位三个步骤。
(1)为密码子所特定的氨基酸tRNA结合到核蛋白体的A位,称为进位。氨基酰tRNA在进位前需要有三种延长因子的作用,即,热不稳定的EF(Unstable temperature,EF)EF-Tu,热稳定的EF(stable temperature EF,EF-Ts)以及依赖GTP的转位因子。EF-Tu首先与GTP结合,然后再与氨基酰tRNA结合成三元复合物,这样的三元复合物才能进入A位。此时GTP水解成GDP,EF-Tu和GDP与结合在A位上的氨基酰tRNA分离(图18-12)。
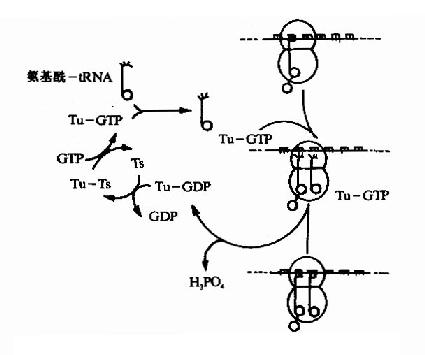
图18-12 原核生物肽链延长因子EFTu与EFTs的作用原理
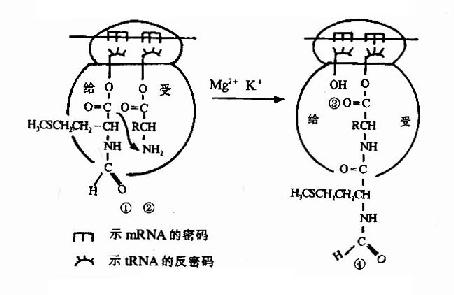
图18-13 肽键的形成
①核蛋白体“给位”上携甲酰蛋氨酰 基(或肽酰)的tRNA
②核蛋白体“受体”上新进入的氨基酰tRNA;
③失去甲酰蛋氨酰基(或肽酰)后,即将从核蛋白体脱落的tRNA;
④接受甲酰蛋氨酰基(或肽酰)后已增长一个氨基酸残基的肽键
(2)转肽--肽键的形成(peptide bond formation)
在70S起始复合物形成过程中,核糖核蛋白体的P位上已结合了起始型甲酰蛋氨酸tRNA,当进位后,P位和A位上各结合了一个氨基酰tRNA,两个氨基酸之间在核糖体转肽酶作用下,P位上的氨基酸提供α-COOH基,与A位上的氨基酸的α-NH2形成肽键,从而使P位上的氨基酸连接到A位氨基酸的氨基上,这就是转肽。转肽后,在A位上形成了一个肽酰tRNA(图18-13)。
(3)移位(Translocation)
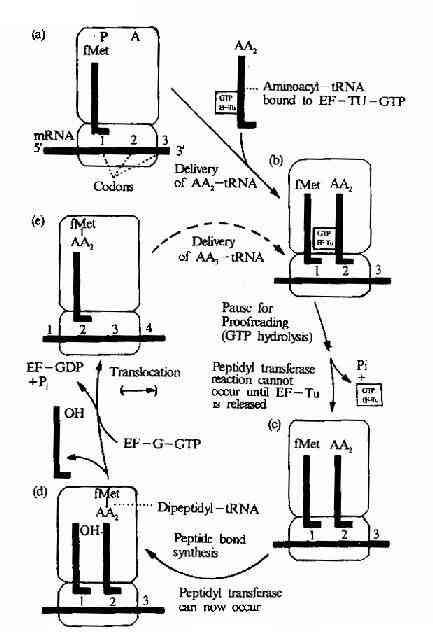
图18-14 Diagram of the elongation process in protein synthesis followinginitation.tRNAs are shown as lue lines;AA,aamicoacyl group.The positioningshown of the EF-Tu-GTP on the tRNA and on the ribosome are arbitraty .Thisdiagram does not make evident why the ribosomal enzyme catalysing the peptidebond synthesis is called peptidyl transferase .However,if you do the next roundof synthesis yourself(see text),you will see that in all subsequent rounds ofsynthesis it is a peptide that is transferred to the incoming aminoacyltRNA-hence the name.
转肽作用发生后,氨基酸都位于A位,P位上无负荷氨基酸的tRNA就此脱落,核蛋白体沿着mRNA向3’端方向移动一组密码子,使得原来结合二肽酰tRNA的A位转变成了P位,而A位空出,可以接受下一个新的氨基酰tRNA进入,移位过程需要EF-2,GTP和Mg2+的参加(图18-14)。
以后,肽链上每增加一个氨基酸残基,即重复上述进位,转肽,移位的步骤,直至所需的长度,实验证明mRNA上的信息阅读是从5’端向3’端进行,而肽链的延伸是从氮基端到羧基端。所以多肽链合成的方向是N端到C端(图18-15)。
|
|
|
| 图18-15 The elongation phase of protein synthesis on a ribosome.The three-step cycle shown is repeated over and over during the synthesos of a protein chain.An aminoacyl-tRNA moleculie binds to the A-site on the rebosome is step l,a new peptide bond is formed in sted 2,and the ribosome moves a distance of three nucleotides along the mRNA chain in step 3,ejecting an old tRNA molecule and "resetting" the ribosome so that the next aminoacyl-tRNA molecule can bind.As indicated in Figure 6-21,the p-site is drawn on the left side of the ribosome,with the A-site on the right. |
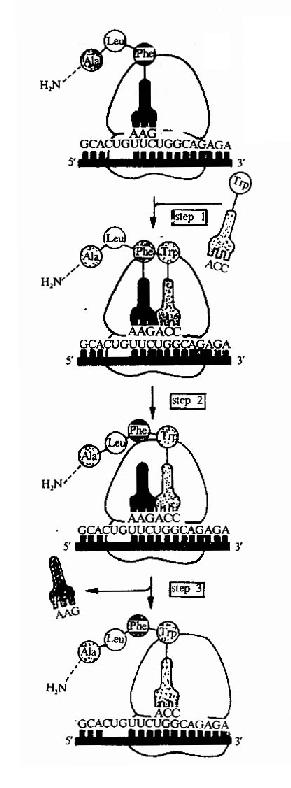
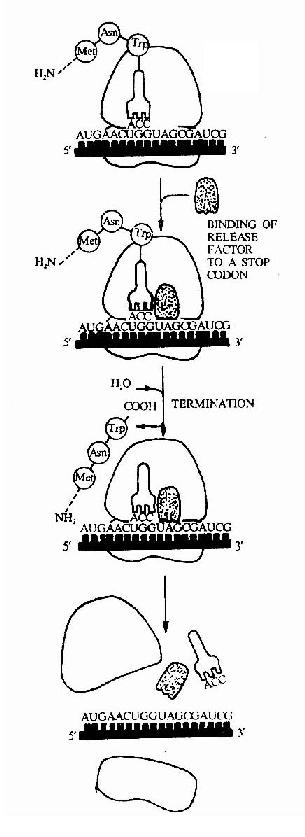 图18-16 The final phase of protein synthesis .The binding of release factor to a stop codon terminates translation.The completed polypeptide is released,and the ribosome dissociates into two separte subunits.
图18-16 The final phase of protein synthesis .The binding of release factor to a stop codon terminates translation.The completed polypeptide is released,and the ribosome dissociates into two separte subunits.(四)翻译的终止及多肽链的释放:
无论原核生物还是真核生物都有三种终止密码子UAG,UAA和UGA。没有一个tRNA能够与终止密码子作用,而是靠特殊的蛋白质因子促成终止作用。这类蛋白质因子叫做释放因子,原核生物有三种释放因子:RF1,RF2t RF3。RF1识别UAA和UAG,RF2识别UAA和UGA。RF3的作用还不明确。真核生物中只有一种释放因子eRF,它可以识别三种终止密码子。
不管原核生物还是真核生物,释放因子都作用于A位点,使转肽酶活性变为水介酶活性,将肽链从结合在核糖体上的tRNA的CCA末凋上水介下来,然后mRNA与核糖体分离,最后一个tRNA脱落,核糖体在IF-3作用下,解离出大、小亚基。解离后的大小亚基又重新参加新的肽链的合成,循环往复,所以多肽链在核糖体上的合成过程又称核糖体循环(ribosome cycle)(图18-16)。
(五)多核糖体循环:
上述只是单个核糖体的翻译过程,事实上在细胞内一条mRNA链上结合着多个核糖体,甚至可多到几百个。蛋白质开始合成时,第一个核糖体在mRNA的起始部位结合,引入第一个蛋氨酸,然后核糖体向mRNA的3’端移动一定距离后,第二个核糖体又在mRNA的起始部位结合,现向前移动一定的距离后,在起始部位又结合第三个核糖体,依次下去,直至终止。两个核糖体之间有一定的长度间隔,每个核糖体都独立完成一条多肽链的合成,所以这种多核糖体可以在一条mRNA链上同时合成多条相同的多肽链,这就大大提高了翻译的效率(图18-17)。
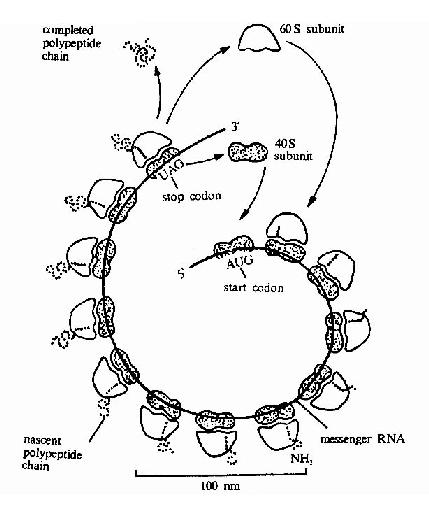
图18-17 A polyribosome.Schematicdrawing showing how a series of ribosomes can simultaneously translate the samemRNA molecule.
多聚核糖体的核糖体个数,与模板mRNA的长度有关,例如血红蛋白的多肽链mNRA编码区有450个核苷酸组成,长约150nm。上面串连有5-6个核糖核蛋白体形成多核糖体。而肌凝蛋白的重链mRNA由5400个核苷酸组成,它由60多个核糖体构成多核糖体完成多肽链的合成。
| 关于“生物化学与分子生物学/蛋白质生物合成过程”的留言: | |
|
目前暂无留言 | |
| 添加留言 | |