组织学/上皮组织的特殊结构
| 医学电子书 >> 《组织学》 >> 上皮组织 >> 被覆上皮 >> 上皮组织的特殊结构 |
| 组织学 |
|
|
|
上皮组织与其功能相适应,在上皮的细胞的各个面常形成不同的特殊结构。这种结构有的由细胞质和细胞膜构成,有的由细胞膜、细胞质和细胞间质共同成。
1.上皮细胞的游离面
(1)细胞衣(cell coat):又称糖衣,为一薄层绒毛状的复合糖,包括糖蛋白、糖脂及蛋白多糖。上皮细胞的游离面细胞衣尤为显着(图2-8),细胞基底面及侧面也有类似细胞衣结构,但不甚明显。细胞衣具有粘着、支持、保护、物质交换及识别等功能。
(2)微绒毛(microvillus):是上皮细胞游离面伸出的细小指状突起,在电镜下才能清楚辨认(图2-8,2-9)。有些上皮细胞微绒毛少,长短不等,排列也不整齐。具有活跃吸收功能的上皮细胞有许多较长的微绒毛,且排列整齐,在高倍镜下可见细胞游离面显纵纹状的纹状缘(straited border)或刷状缘(brush border),这就是密集排列的长微绒毛的光镜图像(图2-3),除上皮细胞外,其它组织的细胞表面也常有微绒毛。
电镜下可见微绒毛表面为细胞膜,内为细胞质。微绒毛直径约0.1μm,长度因细胞种类或细胞生理状态的不同而有很大差别。绒毛轴心的胞质中有许多纵行的微丝(microfilament)。微丝一端附着于微绒毛尖端;另一端下伸到细胞顶部,附着于此部胞质中的终未网(terminal web)(图2-8,2-9)。终末网在吸收功能旺盛的上皮细胞中明显,为顶部胞质中的细丝(filametn)交织成的密网,网与细胞游离面平行,组成网的细丝固着于细胞侧面的中间连接(图2-8,2-9)。微绒毛中的微丝为肌动蛋白丝,终末网中有肌球蛋白。微绒毛大概也以肌丝滑动的方式伸长或缩短。微绒毛显著地扩大了细胞的表面积,参与细胞吸收物质的作用。
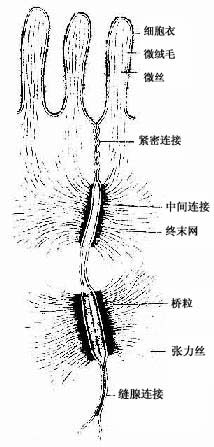
图2-8 单层柱状上皮细胞连接 超微结构模式图
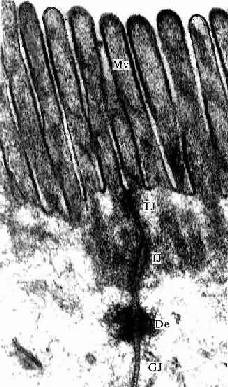
图2-9 小鼠小肠上皮细胞电镜像 ×84000
MV 微绒毛,TJ紧密连接,IJ中间连接,De桥粒,GJ缝隙连接
(上海医科大学电镜室供图)
(3)纤毛(cilium):是细胞游离面伸出的能摆动的较长的突起,比微绒毛粗且长,在光镜下能看见。一个细胞可有几百根纤毛。纤毛长约5-10μm,粗约0.2μm,根部有一个致密颗粒,称基体(basal body)。纤毛具有一定方向节律性摆动的能力。许多纤毛的协调摆动像风吹麦浪起伏,把粘附在上皮表分泌物和颗粒状物质向一定方向推送。例如呼吸道大部分的腔面为有纤毛的上皮,由于纤毛的定向摆动,可把被吸入的灰尘和细菌等排出。
纤毛的内部结构比微绒毛复杂。电镜下可见纤毛表面有细胞膜,内为细胞质,其中有纵向排列的微管。微管的排列有一定规律,中央为2条完整的微管。周围为9组成对的双联微管(图2-10,2-11)。基体的结构与中心粒基本相同,纤毛中的微管与基体的微管相连。
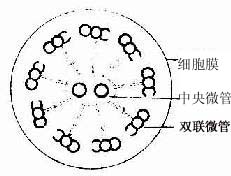
图2-10 纤毛横切面超微结构模式图
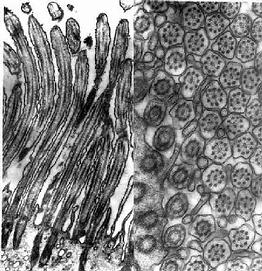
图2-11 大鼠输卵管上皮细胞纤毛电镜像 ×50000
左图 纤毛纵切, 右图 纤毛横切
(上海医科大学电镜室供图)
微管与纤毛的摆有关。有人从纤毛的双联微管中分离出一种具有ATP酶活性的蛋白质,称动力蛋白(dynein)。纤毛的运动可能是此种蛋白质分解ATp 使微管之间产生滑动所致。
2.上皮细胞的侧面在细胞紧密的组织,如上皮组织和其它某些组织,细胞排列密集,细胞间隙很窄,一般宽约15-20nm。细胞间隙中充满相邻细胞的细胞衣,并有少量糖胺多糖和钙离子,有较强的细胞粘着作用。这种粘着物质,在细胞的相邻面间广泛存在。此外,有些细胞的相邻面凹凸不平,互相嵌合,又进一步加强了细胞彼此的结合。细胞间结合更重要的结构,是在细胞相邻面形成特殊构造的细胞连接(cell junction)。上皮细胞间的连接结构发达,而且结构典型。肌细胞之间和神经细胞之间也有较多的细胞连接。结缔组织的细胞较分散,但有些相接触的细胞也有细胞连接。细胞连接由相邻细胞间局部特化的细胞膜、胞质和细胞间隙组成。
(1)紧密连接(tight junction):又称闭锁小带(zonula occludens)。这种连接呈点状、斑状或带状,带状的较典型,常见于单层柱状上皮和单层立方上皮,位于相邻细胞间隙的顶端侧面,呈箍状环绕细胞。在紧密连接的连接区、相邻两细胞的胞膜上有呈网格状的脊,这些脊彼此相对并紧贴在一起,细胞间隙消失。无脊的部分,有10-15nm的间隙(图2-8)。紧密连接除有机械连接作用外,更重要的是封闭细胞顶部的细胞间隙,阻挡细胞外的大分子物质经细胞间隙进入组织内。
(2)中间连接(intermediate junction):又称粘着小带(zonula adherens)。这种连接多为长短不等的带状,位于紧密连接下方,环绕上皮细胞顶部。相邻细胞之间有15-20nm的间隙,间隙中有较致密的丝状物连接相邻细胞的膜。在胞膜的胞质面,附着有薄层致密物质和细丝,细丝参与构成终末网(图2-8)。此种连接在上皮细胞间和心肌细胞间常见。它除有粘着作用外,大概还有保持细胞形状和传递细胞收缩力的作用。
(3)桥粒(desmosome):又称粘着斑(macula adherens)。呈斑状连接,大小不等,位于中间连接的深部,主要存在于上皮细胞间。连接区的细胞间隙宽约20-30nm,其中有低密度的丝状物,间隙中央有一条与细胞膜相平行而致密的中间线,此线由丝状物质交织而成。细胞膜的胞质面有较厚的致密物质构成的附着板,胞质中有许多直径10nm的角蛋白丝(张力丝)(tonofilament)附着于板中,并常折成袢状返回胞质,起固定和支持作用。桥粒是一种很牢固的细胞连接,在易受机械性刺激和磨擦的复层扁平上皮中多见(图2-8)。
在某些上皮细胞的基底面,即与深层结缔组织的相邻面,还可见半桥粒(hemidesmosome)(图2-12)。半桥粒为上皮细胞一侧形成桥粒一半的结构,将上皮细胞固着在基膜上。
(4)缝隙连接(gap junction):又称通讯连接(communication junction)。这种连接呈斑状,位于柱状上皮深部。此处细胞间隙很窄,仅2-3nm,并见相邻两细胞的间隙中有许多间隔大致相等的连接点(图2-8)。冷冻蚀刻复型等方法的研究证,相邻两细胞的胞膜中有许多配布规律的柱状颗粒,每个颗粒直径约7-9nm ,由6个亚单位并合组成,中央有直径约2nm的管腔。相邻两细胞膜中的颗粒彼此相接,管腔也通连,成为细胞间直接交通的管道(图2-13)。在钙离子和其它因素作用下,管道可开放或闭合。这种连接广泛存于胚胎和成体的多种细胞间,可供细胞相互交换某些小分子物质和离子,借以传递化学信息,调节细胞的分化和增殖。此种连接的电阻低,在心肌细胞之间,平滑肌细胞之间和神经细胞之间,可经此处传递电冲动。
以上四种连接,一般只要有两个或两个以上的连接挨在一起,即可称连接复合体(junctional complex)。连接结构的存在和数量常随器官不同发育阶段和功能状态及病理变化而改变。例如,在生精过程事,随着精原细胞的分化,支持细胞间的紧密连接可开放和重建(见男性生殖系统)。

图2-12 半桥粒和基膜超微结构模式图
3.上皮细胞的基底面
(1)基膜(basement membrane):又称基底膜。是上皮基底面与深部结缔组织间的薄膜。不同部位上皮的基膜甚至同一基膜的不同区域,其组成成分有所不同。基膜含有Ⅳ型胶原蛋白、层粘连蛋白和硫酸乙酰肝素蛋白多糖,有的部位基膜含纤维粘连蛋白。电镜下可将基膜分为三层(图2-12):紧贴在上皮细胞基底面的一层为透明板(lamina lucida),为电子致密度低的薄层,厚约10-50nm;其下面为电子致密度高的均质层,称致密板(lamina densa),又称基板,不同部位致密板厚度不等,约为20nm~300nm;第三层为网织板(lamina fibroreticularis),又称网板,位于致密板之下,由网状纤维和基质构成,有时可有少许胶原纤维。基膜厚薄不一,薄者仅由透明板和致密板组成。基膜除有支持和连接作用外,还是半透膜,有利于上皮细胞与深部结缔组织进行物质交换。基膜还能引导上皮细胞移动并影响细胞的分化。
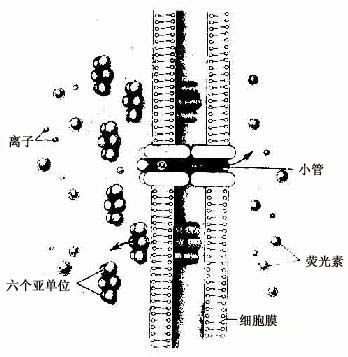
图2-13 缝隙连接超微结构模式图
↑示小分子物质经缝隙连接的小管进入相邻细胞
层粘连蛋白(laminin LN)是基膜中的一种主要成分,是一种大分子糖蛋白,含糖15%-28%,是由一条重链(A链)和2条轻链(B1,B2)构成的不对称的十字形结构,有3条短臂和一条长臂。层粘连蛋白具有与Ⅳ型胶原蛋白、硫酸乙酰肝素、半乳糖脑硫脂及神经节苷脂等分子结合的部位,这对基膜的构成具有重要意义。层粘连蛋白还可与上皮细胞、内皮细胞、神经细胞、肌细胞及多种肿瘤细胞相结合,促进它们粘着在基膜的Ⅳ型胶原蛋白上并铺展开,促进上皮细胞增殖、损伤神经元的存活及轴突生长。层粘连蛋白对保持细胞间的粘着和极性结构以及调节细胞分化皆有重要作用。最近证明,层粘连蛋白亦可增强由抗体或补体介导的巨噬细胞的吞噬功能,增强巨噬细胞对肿瘤细胞的杀伤作用。
(2)质膜内褶(plasma membrane infolding):是上皮细胞基底面的细胞膜折向胞质所形成的许多内褶(图2-14)。质膜内褶的主要作用是扩大细胞基底部的表面积,有利于水和电解质的迅速转运。由于转运过程中需要消耗能量,故在质膜内褶附近的胞质内,含有许多纵行排列的线粒体。
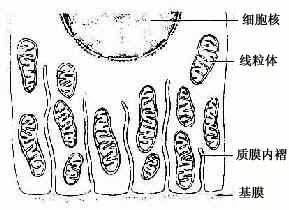
图2-14 上皮细胞基底面质膜内褶
超微结构模式图
| 关于“组织学/上皮组织的特殊结构”的留言: | |
|
目前暂无留言 | |
| 添加留言 | |